大暴发导致“种群崩溃”,历2年不能恢复元气,这对小家鼠种群自身也是不利的,其实是它一次生育量上的失控。怀孕率猛降为0的极端情形却让我们发觉了它深层“秘密”:种群具有“密度-生殖力负反馈”自动调节机制,其种群密度能反向调节生殖力。
高密度下 小家鼠发生生理改变
当年我们通过对北疆小家鼠不同密度年份活体取样检验,结合室内饲养实验,观察到高密度组鼠因拥挤效应,鼠间争斗加剧,内分泌改变。高密度下,小鼠肾上腺皮质肥大,重量增加;幼体胸腺及成体睾丸重量则隨种群密度提高而减轻,呈负相关态势。这反映种群高密度激发肾上腺皮质分泌加强,垂体的生长激素和促性腺激素分泌受抑制。这不仅使幼体生殖器官发育延缓,个体趋小,而且成体睾丸和雌体卵巢亦衰退,睾丸普遍软化、退缩,子宫角变细变薄。
密度负反馈 隨时调节着生殖力
再通过对1970-1980共11年(都是月中旬调查,下文凡述月份即指其中旬)数据作数理统计分析,发现种群密度对雌性生殖力的影响,既表现在不同数量级年份之间,也表现在同一年份的不同增长阶段,负反馈效应都十分强烈。其10月的怀孕率和平均胎仔数,不仅与本期的捕获率极显著负相关,而且与8月的捕获率呈极显著或显著负相关,表明当捕获率达到10%左右时,密度抑制作用即已开始。而负反馈作用于性成熟速度、生殖行为,再到怀孕率、胎仔数等表象上显现,要滞后一二个月。这在生态学上称为“时滞”(time lag)效应。
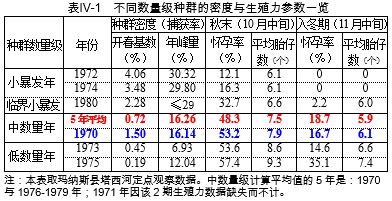 |
相反,对应的两个小暴发后翌年—1973和1975为低数量年,其10月怀孕率为53.6%、57.4%,平均胎仔数为8.6与9.3个,显著高于中数量年同期,11月亦多项偏高,表明是低密度促使生殖力提升。
更有趣的是1980年为临界小暴发年,年峰量大概仅接近29%,其10月与11月的怀孕率和平均胎仔数都比中数量年低得多,却显著高于上述2小暴发年,入冬时怀孕率不归零,足见生殖力受抑程度与种群密度的高低紧密关联。
密度调节与外因促发 鼠种群在各数量级间波动
以上事实表明,北疆小家鼠的密度负反馈调节机制是隨身存在的,种群密度低时会激发繁殖,隨着密度上升则生殖力降低,过高密度就会抑制发育与生殖甚至完全停止。凭藉这个机制,种群通常维持中庸水平。
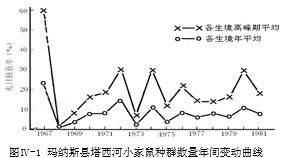 |
1967年那样的大暴发是一次异常,属于种群自身失控。后文将具体分析,由该分析结果可知,该年度种群内外各因素凑成了最优组合,使得越冬鼠存活特多,开春至仲夏各月参加生育鼠甚众,繁殖基数超高,多孕、多胎、多仔,加上发育成熟快速,导致总体增殖过猛过急,而密度走高的“时滞”(从孕育、出生、成长,到入群引发“拥挤”效应,要滞后40~50天)使它来不及反应,数量突然蹿顶再马后炮式负反馈则又过分强烈,终致种群自身崩溃;而种群本来具遗传多型,此时隨着还引发了基因型选择性改变,适于高密度的基因型繁殖力低,所以会延续影响许多世代。这样的失控机率不高,上世纪至少间隔15年或30年才有一次。
种群调节机制有多种学说 小家鼠是多因素综合作用
对动物种群的自然调节机制,历来有各种观点和争论。气候学派认为气候因素起决定性作用,生物学派则突出生物因素的作用。后者认为,竞争、捕食、寄生和疾病等生物因素主要属于“与密度有关因素”,气候基本上是“与密度无关因素”,仅在特定条件下也能成为密度制约性的(例如当隐蔽场所局限,无遮挡保护的个体全部遭恶劣天气杀死,这种死亡是拥挤所致,乃与密度有关);并强调,对种群数量起调节作用的,只能是与密度有关因素。近代更多学者强调内因的作用,形成自动调节学派,按其强调点不同,又可分为行为调节学说,内分泌调节学说和遗传调节学说。从另一角度则有主张,各类生态因素在数量调节上的作用要区别是对变温动物还是恒温动物,变温动物在较大程度上依赖气候因素;恒温动物主要决定于与密度有关的生物因素,种内竞争往往是最主要因素。又有认为,气候稳定并适合生物繁殖的地区,生物因素起主要作用;气候条件差、冬季明显的地区,气候因素会起决定作用。
显然,各学派是从不同的侧面揭示了动物种群自然调节的各项重要因素。我们在北疆小家鼠种群上观察到,实际是多种内因和外因起着综合的作用,不同时期、场合,调节的主导因素不一样。小家鼠每次大暴发、小暴发,显然都同有利的气候和食物条件的保障与促发相关,尤其冬季温暖、厚积雪、多食物的作用甚大;而种群的内部条件,特别是生理生化状态、年龄结构等是否正处于上升态势,则起着基础性作用,若种群内部生理生化正处密度负反馈抑制状态,有利的气候条件就难生效。该种群的密度负反馈机制特别强势,可引发行为改变和内分泌变化,高密度下,个体间攻击行为烈化,肾上腺皮质增大、分泌加强,胸腺萎缩,垂体的生长激素和促性腺激素分泌受抑,终致生殖腺衰退,出生率下降,死亡率升高,种群增长逆转。而大暴发后期种群抗寒力极弱,翌年增殖力低下,则有可能是高密度期产生的子代基因型改变之故。遗传调节学说认为种群具有遗传多型,设想适于高密度的基因型具有高进攻性行为,繁殖力较低,并可能有外迁倾向。依天山北麓农区1967年及伊犂盆地农区1970年小家鼠暴发期间所见,这些迹象确实都有,秋季小家鼠成群结队迁移,夜间啮食儿童指趾和耳鼻,甚至还发生了群鼠围攻小猫的奇闻。高原生物所的研究者在1980-1981年间分5期到玛纳斯现场采取血样,进行北疆小家鼠血红蛋白(Hb)多态型电泳检测,分析其与种群密度的关系,结果观察到该种群有6种Hb多态型,种群密度越高,多态型数也越多,成分就越复杂;Ⅵ型只发现于种群小暴发的1980年10月采集的样本中,其频率很低,并含有泳动最慢的A成分。此型在其他季节未见,很可能属Hb罕见型。这一情况提示,种群密度不同时期产生的个体,确实可能存在基因型的差别,值得进一步探究。
综上所述,可知北疆小家鼠的种群数量调节受着多因素综合作用,其中“密度-生殖力负反馈”內在机制通常起着主导作用,具高灵敏度的自动调节功能,隨身隨时起着调控作用。但时滞效应在特定条件组合下,小概率地可使调控延误,导致大暴发而后种群崩溃。
北疆小家鼠种群崩溃显示动物生育失控对其自身是不利的。因而不少强生殖力鼠种都存在密度-生殖力负反馈自动调节机制,依靠生理机能的调控避免崩溃。人类是智慧生物,更应懂得实行自觉的计划生育和对资源的可持续利用,与大自然和谐共处,永葆种族长青。